連載「博士のホップノート」|第4章 ホップ育種
第4章 ホップ育種
ヨーロッパでのホップ栽培の歴史は1000年以上に及ぶといわれています(Biendlら 2014)。そして今日でも、過去のビールの味わいを「ホップの品種」が教えてくれます。なぜなら、遠い昔からの品種が今なお栽培され続けているからです。代表的なものとして、チェコの Saaz(Saazer とも呼ばれる)、ドイツの Hersbrucker、Spalter、Tettnanger、Hallertau Mittelfrüh、そしてイギリスの Fuggle や Goldings が挙げられます。
品種という概念が確立する以前のホップは、遺伝的には雑多な集団でした。畑ごとに異なる特徴を持ち、さらに近くに雄株の野生ホップがあれば自然交配が起こり、種子がこぼれて新しいホップが芽生えたことでしょう。農家はやがて、畑にさまざまなホップが混在していることに気付き、より栽培しやすく収量の多い株を選び取るようになりました。こうして長い年月をかけて、農業的特性だけでなく、味や香りの点でも優れたものが選び抜かれ、時を超えて受け継がれてきたのが、前述の品種群なのです。
品種名には、その土地の名前が付けられています。末尾の「-er」は「~の」という意味を示し、たとえば Saaz 地方のホップは Saazer、Hersbruck 地域のホップは Hersbrucker となります(-er を省略する場合もありますが、同じ品種です)。一方イギリスでは、Fuggle や Golding といった品種があり、これは農家の名前に由来します(Burgess 1964)。18世紀に IPA に使われたホップの子孫である可能性もあります。
Golding からはその後 Kent Golding、Canterbury Golding など複数の系統が選抜され、総称して Goldings と呼ばれるようになりました。古い時代のホップは、まさに雑多で多様な集団だったのです。
交配育種の始まり
経験則に基づく交配は古くから行われていたようですが(Neve 1991)、19世紀末にメンデルが遺伝の法則を発見したことで、遺伝学を基盤とした科学的な品種改良が始まりました(Biendlら 2014)。1907年には、イギリスの Wye College において Salmon 博士が交配育種を開始します。博士は広い視野を持ち、病害抵抗性や栽培のしやすさ、収量の多さといった農業特性にとどまらず、当初から「品質」にも注力していた点が特徴的です。
Salmon 博士は、高い樹脂含量を持つアメリカのホップと、優れた香りを持つヨーロッパのホップを交配し、両方の特性を兼ね備えた品種を生み出そうとしました(Neve 1991)。当時、育種の現場ではα酸がまだ測定できなかったのか、α酸とは言っていません。「高い樹脂含量」という表現が用いられており、これは肉眼で観察できるルプリンの量を指していたと考えられます。香りについては実際に嗅覚で判断しており、まさに五感を駆使した育種でした。樹脂には α酸が含まれるため、実質的には高 α酸含量を狙った品種改良であったといえます。
1918年には、カナダ・マニトバの野生ホップを雄株として畑に植え、自然交配(Open Pollination、OP とも略されます)を行いました。その交配種子から選抜されたのが Brewer’s Gold と Bullion です。これらの品種は Fuggle や Goldings の約1.5倍もの α酸を持ちます。
Brewer’s Gold の系譜からは、さらに α酸含量の高い Northern Brewer が生まれました。この品種は、当時ドイツで流行していた土壌伝染性カビによる Verticillium 病に抵抗性を持っていたことから、母本としても利用され、Perle が育成されています。
一方アメリカでは、Brewer’s Gold を祖先にもつ高α酸品種が次々に誕生しました。Galena、Nugget、Chinook といった、今や有名な品種群です。今日ではホップの品種数は300近くにのぼるとされますが、高α酸系統のルーツをたどると、Brewer’s Gold か北米の野生ホップに行き着きます。そもそも Brewer’s Gold 自体がカナダの野生ホップ由来でした。
交配育種で生まれた香りの良い品種としてまず挙げられるのは Cascade でしょう。Fuggle の系統を持ち、1972年に誕生しました。クラフトビール人気の原動力になった品種と言っても過言ではありません(富江 2019)。
私自身、クラフトビールの品質に懐疑的だった頃、アメリカで初めて Sierra Nevada Pale Ale を飲みました。とにかく美味しかった。ホップ香が際立ったビールは好きでしたが、度を超えて攻撃的な香味のビールに出会うことも少なくありませんでした。しかし、このビールは違ったのです。強烈でありながら繊細なバランスで非常に飲みやすく、「これが Cascade のビールか」と感動しました。同じ頃、Hallertau Mittelfrüh を用いた Samuel Adams の Boston Lager にも強い感銘を受けました。これならクラフトビールが人気になるのも当然だと納得しました。
Cascade の話が出たので、その育種の経緯に少し触れておきます。Cascade はオレゴン州立大学の Haunold 博士(アメリカ農務省にも所属)が育成した品種です。信じがたいことですが、当初の Cascade は華々しいデビューからは程遠く、普及は困難を極めました。最大のユーザーとなるはずのアメリカの大手ビール会社が、このホップを使おうとしなかったのです。
当時のホップ事情は次のとおりです。ホップは大きく、α酸の高いビターホップと、香りのよいアロマホップに分類されていました。ビターホップについては「α酸を高める」という明確な育種目標が設定できます。しかし問題はアロマホップです。「良いホップの香り」とは何か、「アロマ」とは何か、その定義が不明確だったのです。
当時、チェコの Saaz やドイツの Hallertau Mittelfrüh などが「アロマホップ」として栽培され、大手ビールメーカーに使用されていました。一方、アメリカには相当する品種が存在しませんでした。そこで Haunold 博士は「アメリカのアロマホップを育成する」という挑戦に取り組み、実際に作り出しましたが、その成果は否定されてしまったのです。
1980年、Anheuser-Busch 本社の会議室で熱い議論が交わされました。このクローズド会議の様子は、参加者の一人である鏡ら(1985)によって記録されています。参加者は20名ほどで、Haunold 博士と育種部門スタッフ、アメリカの大手ビール会社3社、日本1社が出席していました。議題は「アロマホップとは何か」「その成分指標は何か」であり、品種改良においてホップを選抜するための基準を模索していたのです。
1980年代前半、大手ビールメーカーが生産するビールの種類は決して多くなく、主力はラガービールでした。そうした状況下で、各社の意向に合致し、共通して「アロマホップ」と呼べる品種を求めていたのです。限られた優良品種を用い、それぞれのメーカーがビールを仕込むという時代でした。多様化した現代の視点からすると、この議論は画一的で無理のあるものに思えますし、「アロマホップとは何か」という問いに唯一の答えがあるとも考えにくいでしょう。しかし当時、世界のホップ研究者たちはその答えを求め、分析に注力していたのです。
アロマホップの指標は後に示されました。具体的には、β酸画分/α酸が 1.6 以上、cohumulone 比率が 25% 以下、humulene/β-caryophyllene が 3.0 以上といった基準です。これらの指標は世界の主要な育種機関に受け入れられ、ホップ選抜の基準となっていきました。苦味の章で既に触れましたが、ここで再び cohumulone 比率が登場します。後ほど改めて深掘りすることにします。
ここで注意したいのは、この基準はあくまで「ビール」ではなく「ホップ」に対する指標である点です。品種改良の現場では膨大な数のホップを扱うため、その都度ビールを仕込むことはできません。ビール化は最終評価の段階で行われるため、上記の数値はあくまで代替指標として利用されていたのです。言い換えれば、経験則に基づく基準でした。
しかし現在では、この指標は必ずしも有効ではありません。Cascade や Citra のように多様で特徴的な香気を持つ品種は、この範囲に収まらないからです。
品種の話に戻りましょう。古くからの知り合いである Gene Probasco が育種したアメリカの Citra は、Hallertau Mittelfrüh の系譜を受け継いでいます。同じくアメリカの Centennial は Brewer’s Gold を起源とし、個性的な香りを特徴としています。ニュージーランドの Nelson Sauvin はアメリカの Cluster の系統に連なり、そのルーツをたどるとイギリスのホップに行き着きます。Burgess(1964)によれば、イギリスのホップは 1629 年に移民とともに新大陸へ持ち込まれました。当時から現在に至るまで栽培が続けられている Kent 地方を起源とする Fuggle や Goldings に関連する系譜と考えられるでしょう。
このように品種の背景を俯瞰すると、ホップはまさにグローバルな存在であり、多様な遺伝的背景が今日の多様性を支えていることが分かります。
現在のホップ新品種開発の状況については第6章でまとめますが、新品種は今後も世界各地で次々と登場するでしょう。問題は、クラフトビール市場が求めるような新たな香気を備えたホップを狙って育成できるのか、という点にあります。以降では、遺伝学の観点から苦味や香りについて考察していきます。
雄ホップと交配育種
ホップの雌雄は、基本的に異なる個体です。雄ホップ、雌ホップがあるのです。基本と断ったのは、雄雌の両性があるからです。雌の花の中に一部、雄花を付けるのです。間性と呼んでいます。
ホップの性別を決めるのは遺伝子であり、人間と同じように X と Y の性染色体を持っています。ところが、性染色体以外の常染色体上の遺伝子も関与していると考えられています(Neve 1991)。X 染色体と常染色体にある複数の遺伝子の比によって、雌、雌寄りの間性(着花数:雌花が雄花より多い)、雄寄りの間性(着花数:雄花が雌花より多い)、そして雄へと分離します。Y 染色体は、生殖能力を持つ花粉を形成するために必須です。
栽培されるホップはすべて雌株です。雄株が存在するのは、育種研究機関で遺伝資源として保存されている場合か、野生ホップとして自生している場合に限られます。
育種は、交配によって遺伝子の組み合わせを変えたり、遺伝子そのものに変異を起こさせたりすることから始まります。ここでは一般的な手法である交配育種について説明します。
まず花粉を集めます。雄花は直径1cmに満たない小さな粒で、房状に多数がつきます(図4-1)。5枚の花被が開くと開花であり、花粉をため込んだ葯が現れます。この状態の雄花を収穫し、乾燥させます。軽く揉むと黄色の粉が舞いますが、これが花粉です。
次は受粉です。雌の本来の花である毛花(図4-1)のめしべに花粉を付けます。耳かきの綿毛を使って一花ずつ受粉させたり、雌花の枝に紙袋をかけ、その中に花粉を吹き込む方法もあります。受粉・受精が成功した毬花は内苞が大きく成長するため、一目で判別できます。緑色の毬花が黄色みを帯びたら収穫し、自然乾燥させます。
種子はゴマ粒のように小さく、100粒で約1g程度です。乾燥した毬花を両手で揉むと種子がこぼれ落ちますが、最終的には指先の感覚を頼りに一粒ずつ回収します。採取した種子は少なくとも1か月間冷蔵庫で保存し、「冬」を与えます。冬を経験しないと春を感知せず、発芽しないのです。ホップが四季のある環境に適応し進化してきた証拠といえるでしょう。

発芽したホップは蔓をぐんぐん伸ばし、枝葉を茂らせ、地下では根を張り栄養を蓄えて株へと成長します。これを実生苗と呼び、翌年に定位置へ植え直します。個体ごとに遺伝子が異なるため特性もさまざまです。その特性を見極めるため、隣同士が絡まないよう間隔をあけて植え付けます。
ただし、植え替え1年目は本来の姿を示さないことが多く、2年目になってようやく見極めが始まります。収量性や耐病虫害性といった農業特性が良好な個体を選び出す作業、これを選抜と呼びます。加えて、毬花の香りや化学成分を確認し、合格したものを翌年から増殖します。
増殖は地下に埋もれた蔓の部分を切り分けることで行います。地上部の蔓は冬に枯れますが、十分に土がかかっている地下部は乾燥に耐えて生きており、その部分を利用します。こうして1個体から群をつくり、再評価を行います。収穫量も増えるので実際にビールを仕込んで評価します。
結果はがっかりするか、飛び上がって喜ぶか、まさに運次第です。ここで「運」と言った理由については、後半で説明します。
遺伝の基礎
以降でホップの品種改良や遺伝を説明するにあたり、いくつかの用語解説と遺伝学の復習が必要になります。触り程度ですが、少々お付き合いください。
細胞のゲノムは基本的に対になって構成されています。このような生物を 二倍体 と呼びます(もちろん三倍体、四倍体も存在しますが、ここでは二倍体を前提に説明します)。つまり、一対の染色体で構成されているということです。
生殖細胞では減数分裂によって染色体が半分になり、ゲノムも半数体となります。その後、受精によって再び対の二倍体、すなわち元のゲノム構成に戻ります。対である以上、一方に遺伝子があれば、もう一方にも対応する遺伝子が存在します。そして遺伝子の間には、優劣といった力関係も存在するのです。
質的形質
花の色のように赤、ピンク、黄といったように、はっきり区別できる不連続な形質を質的形質と呼びます。この質的形質の遺伝には、メンデルの法則をあてはめて考えることができます。
色を決める遺伝子を A、その対になる遺伝子を a とします。どのような組み合わせの遺伝子を持つかを遺伝子型と呼びます。同じ遺伝子を持つ場合はホモ型(AA、aa)、異なる場合はヘテロ型(Aa)と呼びます。そして花びらの色など、実際に現れる性質を表現型と呼びます。
遺伝子型がヘテロ型同士、つまり Aa × Aa で交配すると、子世代の遺伝子型は AA、Aa、aa の3種類に分離し、その比率は 1:2:1 となります。
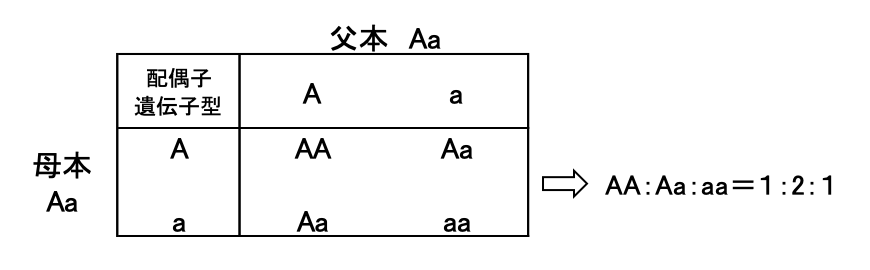
遺伝子型 AA を赤、Aa をピンク、aa を黄と表現型に対応させると、赤:ピンク:黄の比率は 1:2:1 になります。このように AA と Aa が異なる表現型(赤とピンク)を示す場合、A は 不完全優性 の主働遺伝子と呼ばれます。
一方、AA と Aa が同じ表現型(赤)を示す場合には、A は 完全優性 の主働遺伝子と呼ばれます。このときピンクの表現型は消失し、赤:黄の比率は 3:1 となります。なお、a は劣性遺伝子と呼ばれます。
量的形質
ホップの α酸含量や収量は、交配によってさまざまな値を示す子どもの集団が得られます。これを 2 次元の棒グラフにしてみましょう。横軸には α酸を 1%、2%、3%といった幅で区切って取り、縦軸にはそれぞれの範囲に該当する子どもの数を示します。これを度数分布と呼びます。多少の歪みはあるかもしれませんが、山の頂点から緩やかな稜線を描くグラフが現れます(図4-2)。
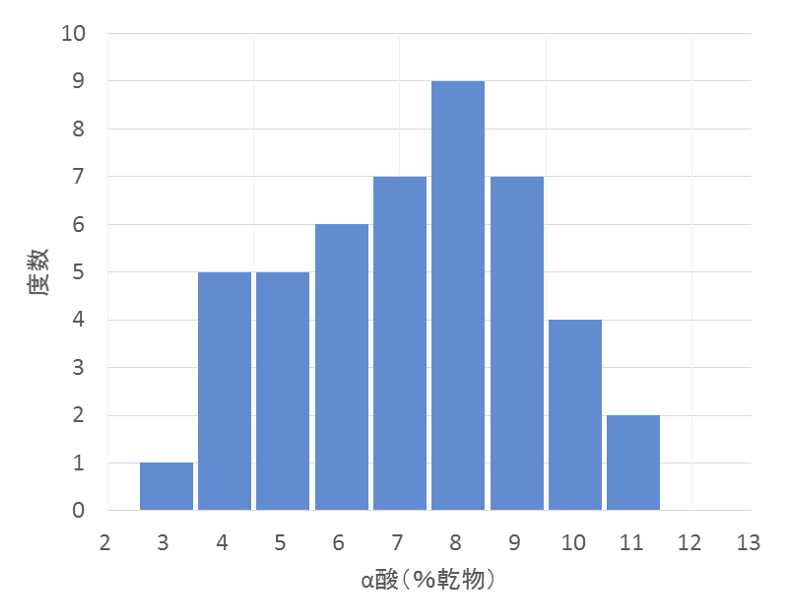
このような分布を示す形質は量的形質と呼ばれます。量的形質を決定するのは、作用の小さい多数の遺伝子が関与しているとされ、その遺伝子の数によって表現型の値が決まります。量的形質に関わる遺伝子は量的遺伝子、あるいはポリジーンと呼ばれます。量的遺伝子は数が多いため、一つ一つを個別に取り扱うのは煩雑であり、統計的に「量」として扱うのが一般的です。これに対し、質的形質は個々の遺伝子単位で取り扱います。
ホップの遺伝学
ホップの代表的成分について、その遺伝の様子を見てみましょう。ここでは、特定の交配から得られた子孫を用いて解析します。図4-2は、ひとつの交配から得られた集団、すなわちすべて雌株で構成される姉妹集団におけるα酸の分離の様子(度数分布)を示したものです。子株ごとに両親から受け継ぐα酸関連遺伝子の数が異なるため、α酸含量の多寡が生じ、連続的な分布を示します。精油成分であるβ-caryophyllene(図4-3)、humulene(図4-4)についても同様で、これらの成分はいずれも明らかに量的形質です。
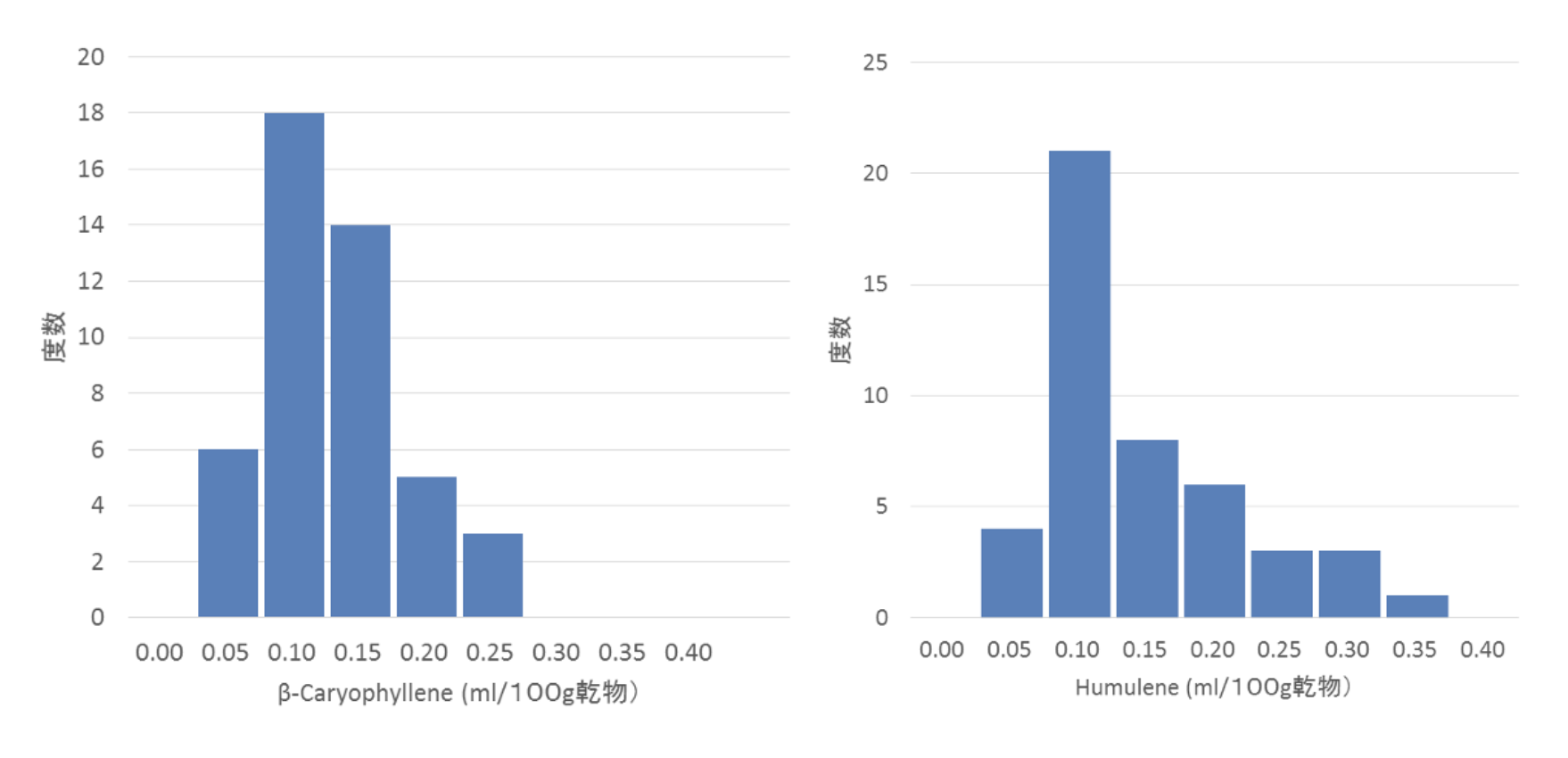
左:図4-3 β-caryohylleneの分離(度数分布)、右:図4-4 Humuleneの分離(度数分布)
遺伝の様子を数値化します。できるだけ多様な遺伝的背景を持つ(母親も父親も異なる)46のホップ個体を複数年栽培し、α酸、精油成分を調べました。露地での栽培なので、気温、降水量、日照等、取り巻く環境の影響を受けます。2年分のデータから「Repeatability」(Falconer1989)という統計量を算出しました(ノート4-1)。厳密な比喩ではありませんが、成分量などの表現型値が2年間の間でどの程度変わらずに安定しているかを示す指標です。値が1に近ければ、環境の影響を受けにくい形質といえます。表4-1にα酸、cohumulone(以下Coh)、β-caryophyllene(以下Car)、α-humulene(以下Hum)の値を示しましたが、いずれも0.8前後と高い値を示しました。HumとCarの比率(Hum/Car)では0.9台です。それはそうでしょう。例えばα酸が10%の品種が、ある年は2%になったり18%になったりしません。大きく増減はしないでしょうが、毎年同じ値というわけにはいきません。例えば8から12%と多少はばらつきます。このようなばらつきが環境の影響と言えます。
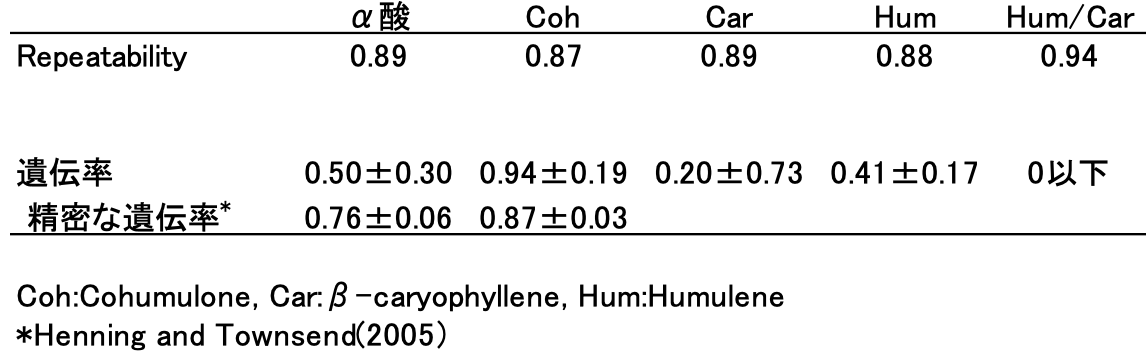
次に「遺伝率」という統計量を算出します(ノート4-1)。どの程度、親から子へ形質が遺伝するのか、言い換えれば遺伝的要因がどの程度関与しているのかを調べるわけです。ただし、実験計画の設計によって計算方法は様々であり、環境と遺伝の相互作用など、純粋な遺伝的要因以外の情報も含まれてしまいます。今回、11種類の母親ホップに交配して得られた子どもが、母親とどれほど似ているかを調べました。事情により1年分の測定データのみを用いたため、得られた値はあくまでラフなものです。その結果、α酸は0.5、Carは0.2、Humは0.4といずれも低めの値となりました。試験を実施した1994年は雨不足とダニの発生が重なり、健全なホップの成長環境ではなかったため、親子間の遺伝的な関係性を上回る環境要因の影響が加わり、低めに推定されたものと思われます。なお、α酸についてはアメリカ・オレゴン州立大学のHenningとTownsend(2005)が大規模調査を行い、より精緻な値として0.76を報告しています。
興味深いのはCohumuloneの遺伝です。図4-4に示すように分離の様子はα酸同様に連続分布しているように見えますが、一か所、矢印の部分で分布が分断されているように見えました。他の集団でも同様の分断が観察されています。これは単純な量的形質ではなく、主働遺伝子が関与している可能性があるのではないかと考えました。そこで、両親がヘテロ接合体である場合を想定し、理論分離比と観察分離比の一致度をχ²(カイ二乗)検定で確認しました。
仮に、Aを完全優性でCohを35%以上に高める遺伝子とすると、Aa同士の交配では子どもはAA、Aa、aaに分離します。Aが完全優性であるため表現型はAA=Aaとなり、Cohが高い個体(AA+Aa)と低い個体(aa)の比率は理論上3:1となります。一方、実測値は36:10でした。理論値は34.5:11.5ですので、両者の分離比は確率0.5〜0.8の範囲で有意に一致しました(表4-2)。
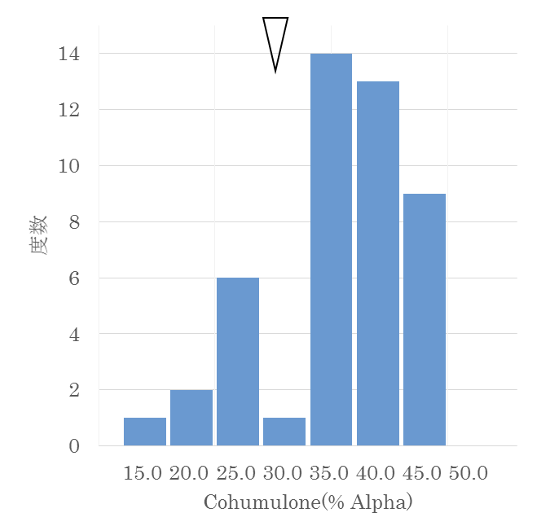
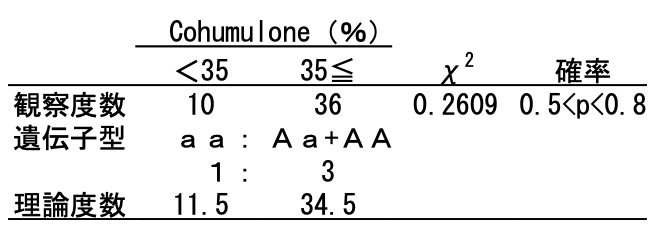
表4-1のCohのRepeatabilityと遺伝率を改めて見てみましょう。前者は0.78、後者は0.94と非常に高い値を示しています。遺伝率のデータを得た年は環境条件が厳しかったにもかかわらず、この数値です。量的形質としては異例に高く、主働遺伝子の存在を支持する結果と考えられます。後にHenningとTownsend(2005)も0.87という高い遺伝率を報告しています。
1999年、ポーランドで開催されたホップ専門学会でこの研究を発表しました。この学会には世界中から研究者が集まります。品種改良、化学分析、栽培、昆虫や病害などの専門家が一堂に会しますが、ホップ研究者は世界的に見ても多くはないため、2年に一度顔を合わせれば互いに知り合いになってしまうほどの規模です。
私は「Coh比率の遺伝には完全優性の主働遺伝子が関与している」と自信を持って発表しました。しかし、イギリスでホップの品種改良を進めていたPeter Darbyは、より大規模な調査に基づき「Coh比率は連続分布し、量的形質の遺伝様式が当てはまる」と発表しました。発表後、直接Peterに「分布が分断するような不連続な結果は見られなかったのか」と尋ねましたが、答えはNo。私の調査は46個体という小規模集団だったため、偶然そのように見えてしまった可能性がある、と感じ自信を失ってしまいました。
ところがその2年後、サッポロビールの岡田さんから「Coh比率には主働遺伝子が存在する。あの発表は正しかった」との言葉をいただきました。彼らはホップの遺伝子地図(Koieら 2005、ノート4-1 連鎖地図)を作成しようとしており、その過程でCoh比率を明確に高・低に分ける遺伝子の存在を突き止めていたのです(Okada 2007)。さらにMcAdamら(2013)は国際協力のもと精緻な遺伝子地図を完成させ、ここでもCoh比率に強力な主働遺伝子を検出しました。
以上を総合すると、Coh比率の遺伝には「主働遺伝子によって高い・低いに大きく分けられる」と同時に、「ポリジーンによって比率が微調整される」と結論づけるのが妥当のようです。主働遺伝子が劣性であっても、ポリジーンが十分に作用すれば、Coh比率の高いホップを生み出すことも可能になります。
Pazakら(2010)は各地の野生ホップのCoh比率を調査しました。表4-3にその一部を抜粋しています。各データは平均値だけでなく標準偏差も示されているため、ばらつきの程度も確認できます。カナダやアメリカの北米集団ではCoh比率が50%台と高く、ヨーロッパ集団および黒海・カスピ海周辺のコーカサス地方集団では20%台と低い値を示しました。しかも標準偏差はいずれも十分に小さく、明確に両地域の野生ホップが「Coh比率の高い集団」と「低い集団」として分化していることが分かります。
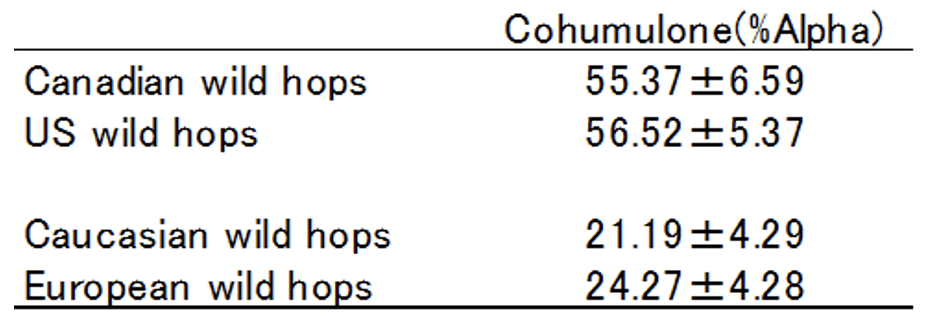
遺伝的分化は、遺伝的交流が遮断され、それぞれが独立した進化を経ることで生じます。ホップの進化の過程については次章で解説しますが、Coh比率が高いという形質は北米集団に見られることをPatzakら(2010)が示しており、この主働遺伝子の起源は北米にあると考えられます。ヨーロッパで見られるのは、人為的に持ち込まれた結果です。Salmon博士が20世紀初頭に交配に用いたのはカナダの野生ホップであり、そこから育成されたBrewer’s GoldやBullionを通じてヨーロッパのホップはCoh比率が高くなり、やがて1970年代の「Cohumulone論争」へとつながっていきます。
遺伝的分化は、遺伝的交流が遮断され、それぞれが独立した進化を経ることで生じます。ホップの進化の過程については次章で解説しますが、Coh比率が高いという形質は北米集団に見られることをPatzakら(2010)が示しており、この主働遺伝子の起源は北米にあると考えられます。ヨーロッパで見られるのは、人為的に持ち込まれた結果です。Salmon博士が20世紀初頭に交配に用いたのはカナダの野生ホップであり、そこから育成されたBrewer’s GoldやBullionを通じてヨーロッパのホップはCoh比率が高くなり、やがて1970年代の「Cohumulone論争」へとつながっていきます。
Cohumulone 論争の考察
苦味の章で触れた、いまだ決着のついていないCohumulone(以下、Coh)由来の苦味の質についてです。この論争で双方が正しいとするとどうなるでしょうか。だからこそ決着がついていないのです。
イソ化したiso-Cohそのものの味については、収れん味などネガティブな表現は報告されていません(Hughes and Simpson 1996、Hughes 2000、Schönberger and Kostelecky 2011)。ベルリンの著名な研究者も同様の見解を示しており、本質はCohそのものではないように思われます。ホップの品種によって苦味の質には良し悪しがあり、悪いとされる方にCoh比率の高い品種が多い(すべてではない)ため、そのように見えているのではないかと考えるようになりました。つまり、相関して「悪さ」をする形質が存在する可能性があるのです。
実際、McAdam(2013)はポリフェノールの一種であるxanthohumolとCohの間にr=0.87という高い表現型相関を報告しています。xanthohumolはホップ特有の成分で、多様な薬理作用が報告されている注目化合物です。熱により異性化し、isoxanthohumolとしてビール中に微量溶け込みますが、味への寄与は低いと考えられています(Biendlら2014)。
さらに、xanthohumolとCoh比率に関わる遺伝子は同じ連鎖グループに属していました。すなわち、遺伝的にも相関しているのです。もしCoh比率に作用する主働遺伝子に、苦味質を悪化させる遺伝子が連鎖しているとすれば、苦味の質の悪さが一見Coh比率と連動しているように見えるのも理解できます。時に連鎖が切れることで例外が生じるはずです。さて、真相はどうなのでしょうか。
1970年代前半に始まったCohumulone論争は、その後のホップ育種に大きな影響を与えました。Cascadeに端を発した「アロマホップ」の定義にもCohが登場するように、Coh比率を低下させる育種が強力に進められたのです。是非が明確でなくとも、苦味質悪化の懸念やリスクを避けた結果でした。そして、決着がついていないために、その影響はいまも残っています。
ブルワー向けのホップ品種カタログには、品種ごとの香りのキーワードや成分組成、α酸含量などの特徴が明記されていますが、Coh比率もほぼ記載されています。テルペンの量や比率が掲載されることもあります。これらの記載は、まさに過去の「いきさつ」が反映されたものなのです。
新ホップ品種MURAKAMI SEVENのいきさつ
この章で使用した表や図は、1991年に交配して得られたホップの姉妹集団のデータです。新品種 MURAKAMI SEVEN は2016年、新しいビール商品とともに突如登場しましたが、その起源はこの集団にあります。
母親となったホップは、青い松ヤニ様の香りが特徴で、とても良い香りとは言えませんでした。しかし、毬花が鈴なりに密集して付くため交配が容易であり、複数の交配を要する遺伝研究には好都合の母本でした。本来はホップの遺伝を調べるための材料だったのです。その後、ビールに仕込んだ際にどのような成分になり、どんな香味を示すのかも調べました。すると、素晴らしく良い香りを持つホップが見つかり、育ち姿も美しく農業特性も十分満たした理想的な姿を示しました。これなら農家も受け入れてくれるはずと考えられました。普及は岩手県遠野市を中心に始まり、2024年からは秋田県横手市でも本格的に栽培が始まります。
育種用の畑は岩手県奥州市江刺にあり、一列に植えた畝の7番目にあったため「江刺7号」と呼ばれるようになりました。キリン社内での評判も良く、2016年には東京・代官山のスプリングバレーブルワリーで少量ながらビールとして発売されることになりました。品種名をどうするかと聞かれた際、私は「任せるよ、自分はセンスが無いから」と答えてしまいました。本当に、うかつでした。まさかの「MURAKAMI SEVEN」。知った時にはすでに手遅れで、印刷物や販促品はすべて揃っていました。
名付けたのは、スプリングバレーブルワリー初代社長の和田さん。酎ハイで数々のヒット商品を生み出した凄腕のマーケッターです。そしてビール造りを支えたのが、ビール全商品の味や香りを決めるトップ中のトップ、マスターブルワーの田山さん。この二人が組んだのですから、もう誰にも止められなかったのです。
そもそもホップの新品種は、具体的な商品が想定される前に、あらかじめ増殖しておく段取りが必要です。商品が決まった時点でまとまった量のホップが用意されていなければ話は進みません。「これから増殖して、使えるのは3年後です」では通用しないのです。このリスクをヘッジするのは研究所の役目です。
2011年の東日本大震災がきっかけでした。甚大な被害を受けた方々に何ができるかと考えたとき、やはりホップしかありませんでした。新しいホップ品種で新しいビールをつくり、届けたい。少しでも喜んでいただきたい。そう思って相談したのが田山さんでした。当時、研究所の副所長の職にあり、即決してくださいました。
ホップの増殖は容易ではありません。最初はわずか1株から始め、1年で10倍にしか増えません。何年もかけて収穫量を積み上げていきました。その間に会社にはクラフトビール部門が立ち上がり、スプリングバレーブルワリーが誕生し、そこで初めて商品化されたのです。今では通年生産の商品にも使えるほどの栽培面積と生産量に達しています。
今年のMURAKAMI SEVENの出来はどうかと、畑を歩き回っています。当たり前ですが、今年もMURAKAMI SEVENらしいホップが収穫されるでしょう。MURAKAMI SEVENはいつもMURAKAMI SEVENなのです。アメリカのCascadeを栽培している人もいますが、日本で栽培してもCascadeはやはりCascadeです。この当たり前の事実は、なぜなのか。ホップ香気の章の最後に触れた疑問です。少しだけ自論を展開してみます。ただし、結論には至っておりませんので、あしからず。
ホップ品種のアイデンティティーとは
MURAKAMI SEVEN の特徴はイチジク、マスカット、抹茶様など、様々に語られますが、どのような香気成分が関わっているのかはまだ分かっていません。含硫成分の 4-methyl-4-sulfanylpentan-2-one が見つかる可能性もあります。いずれ特徴的な香気成分が特定されると思いますが、その成分量は繰り返しになりますが栽培環境によって変動します。
Repeatability が 1.0 になることはあり得ません。量が変動すれば、その特徴香気の強度も変わります。Henning ら(1997)によれば、精油総含量の遺伝率は 0.12 とかなり低く、個々の香気成分量は大きく変動することを示しています。
「一番搾り とれたてホップ」は、岩手県遠野産のホップを乾燥せずに使用します。その年のホップの品質が前面に出るため、IBUKI の特徴が強い年もあれば弱い年もあります。実際に Linalool の増減が確認されました。ただし、強くても弱くても IBUKI は IBUKI です。それを決めるのは香気成分の量ではないようです。
では次に、香気成分の「比率」が関わっているのではないか、ということになります。具体例で説明しましょう。表 4-1 の Repeatability における β-caryophyllene(以下 Car)、α-humulene(以下 Hum)の値と、Hum と Car の比率(Hum/Car)を比べてみます。Car と Hum はいずれも 0.8 台ですが、Hum/Car は 0.92 と著しく高いのです。この比の遺伝的分離を示したのが図 4-6 です。
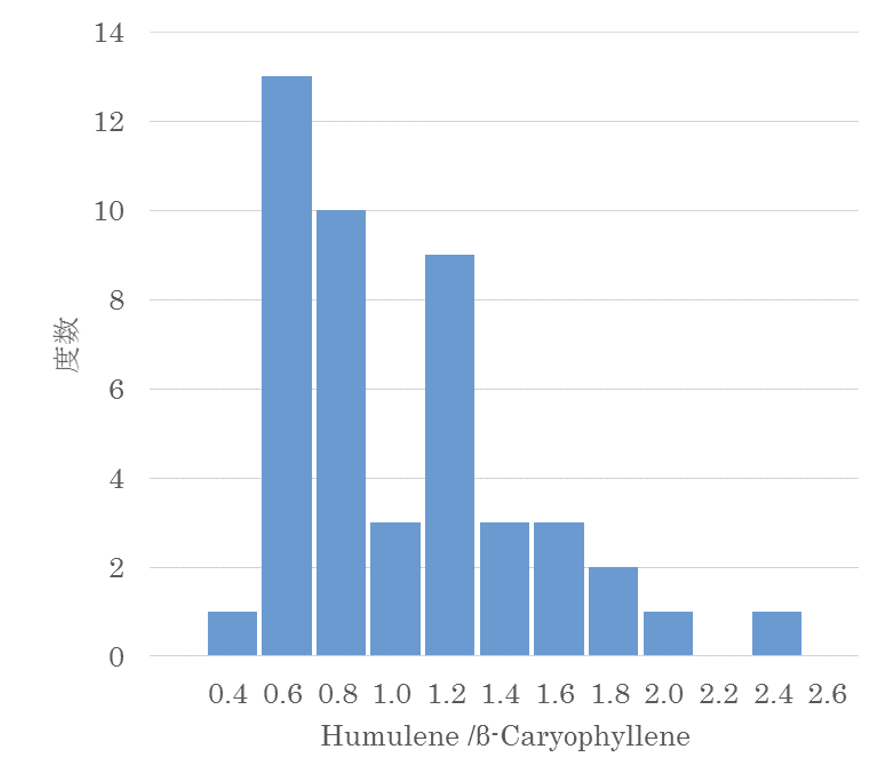
図4-6 α-humulene/ β-caryophylleneの分離(度数分布)
凸凹があり連続分布ではありません。Car と Hum の分離は連続分布していますが、有意な遺伝相関はなく、互いに独立していました。また Hum/Car は遺伝率(表4-1)も計算できませんでした。つまり Hum/Car は遺伝する形質ではなく、Hum と Car がランダムに変動し、その偶然によって生じた形質なのです。
Repeatability で言えば、Car や Hum それぞれの量よりも、Hum/Car 比率の方が高い値を示しています。ところが Wang ら(2008)によれば、Hum と Car の比率を決める酵素が存在するといいます。主働遺伝子があるということであり、これまでの「量的形質としての取り扱い」や「互いに遺伝相関を持たず任意の値をとる」との説明とは矛盾します。
テルペン類の合成経路は複雑です。ひとつの酵素が複数のテルペン類を合成したり、生成されたテルペンが最終産物ではなく、その後に酸化など様々な修飾を受けることもあります(飯島 2014)。Hum と Car は最終成分ではなく、その先の成分の基質にもなっています。例えるなら、本流の川から下流に向かって支流に分かれ、さらに細かな支流へと分かれるイメージです。その分岐は酵素によって決まり、分岐の数だけ酵素が存在するのです。多数の遺伝子(共優性)が関与しているために量的形質のように見えますが、本質的には主働遺伝子が比率を決めており、いったん決まると環境の影響をほとんど受けない頑固な形質となります。このような成分比こそが、ホップ品種の性格やアイデンティティーを強く規定していると考えた方がよいかもしれません。
ただし、成分の比率に注目すると、検証は容易ではありません。どの成分の比を見ればよいのか、比をとる成分の組み合わせは膨大になります。さらに、分析対象とする成分の選択も慎重に行う必要があります。品種間の差から候補を選ぼうとしても、比を構成する成分自体が品種によって異なる可能性があるため、品種間差を手掛かりにはできません。
そこで挑戦したのが、私自身も関わった Furukawa ら(2012)の研究でした。 遺伝的に遠い品種ではなく、遺伝的に近縁の交配集団を用いれば、香気の特徴は連続的に分離してくるはずです。この章で繰り返し登場する姉妹集団のうち 23 個体を用いてビールを醸造しました。その結果、フルーティーな香り、汗様の香り、ココナッツ様の香りと特徴が分かれました。これは、母親や父親から受け継いだ遺伝子の違いによる、遺伝的な差異です。
成分の選択には Repeatability を用い、8 成分まで絞り込みました。香りと成分の間にそれらしい関係も見えましたが、どうにも決定打に欠けました。成分比も調べましたが「それらしい」との結論にとどまりました。比率の変化と香りの変化が連続的ではなく、例えばココナッツ様、汗様、フルーツ様が順番に現れるという秩序性はなかったのです。香りの質を決める法則は見いだせませんでした。
類似性のある姉妹集団でも核心が得られないのであれば、準同質遺伝系統を作るしかありません。同じ親を繰り返し交配し(戻し交配、back cross)、何世代も重ねることで親に近い DNA を持ちながら、わずかに特徴が異なる集団を作ります。その小さな差で香気が大きく変われば、核心に近づけるはずです。ただし、この実験は何年もかかり、香気の違いが現れるかは運に左右されるため、必ずしも成立しないかもしれません(なお、ウルトラC級の実験系についてはノート4-2を参照ください)。
仮に成分比率の核心が分かっても、それが偶然の産物であるならば、育種でのコントロールは容易ではありません。「良い香りのホップがほしい」としても、明確な方向性は示せないのです。さらに、そこに整った農業特性が同時に備わる確率はおそらく低いでしょう。品質と農業特性を両立させるには、地道な努力と時間、そして人の力が必要です。
しかし裏を返せば、偶然の産物だからこそ、新たな香気を生み出す可能性があるのです。実際、世界のホップ品種数は急速に増えています。未知の成分比率が新しい香気をもたらしているのです。新品種の誕生は、まだまだ続いていくでしょう。
ホップのテロワール
この章の最後は、栽培環境と遺伝子の共同作業についてのお話です。
よくある質問のひとつに「ホップの品質と産地の関係」があります。生ホップを使用する「とれたてホップ一番搾り」はラベルに「遠野産ホップ使用」と銘打っているため、遠野のホップは他の産地と比べて何が違うのか、という内容です。答えは「違いはありません」。どの産地のホップも気候の影響を受けて品質は変動しますが、その変動の範囲にすべてのホップが収まってしまうからです。これが一般的な返答になります。
ホップ品種 IBUKI では、産地間で大きな違いは感じられません。しかしホップのテロワールは確かに存在します。栽培環境と遺伝子の安定した相互作用があるからです。
「テロワール」という言葉の定義には微妙な差異がありますが、作物の場合には、その栽培環境(土壌や気象など)や栽培方法、そしてその場所に起因する人的要因の相互作用によって生み出される品質、と解釈できます。
ホップの場合、産地によって栽培方法に違いがあります。分かりやすい例が堆肥の種類です。秋田県大館市比内では比内鶏の飼育が盛んなため、鶏糞が最も入手しやすい堆肥です。牛糞、豚糞、鶏糞と有機質肥料が異なれば成分も変わり、それに合わせて化成肥料の施し方も変わってきます。また、長年の経験則によって作業方法も異なります。
例えば豪雪地帯である秋田県横手市では、冬の間ホップの株の頭は外に露出したままです。一方、遠野では冬の寒さや乾燥から株を守るために、しっかりと土を被せる必要があります。ここに大きな違いがあります。横手ではメートル単位で雪が積もるため土を寄せる必要がなく、春の雪解けとともに自然にホップが萌芽してくるそうです。
テロワールを少し科学的に考えてみましょう。ある特定の環境によって遺伝的な特徴がさらに際立つ場合です。
Falconer(1989)によれば、表現型値の分散(Vp:ノート4-1参照)には、遺伝子型の分散(VG)と環境の分散(VE)に加えて、遺伝子型と環境の共分散(COVGE)が含まれます。数式で表すと、
Vp=VG + VE + 2COVGE
となります。
この遺伝子型と環境の共分散とは、簡単に言えば「相性」のようなものです。品種によって相性が良い場合もあれば悪い場合もあり、その結果、共分散が発生したりしなかったりするのです。
Holle ら(2021)は、アメリカのホップ品種 Amarillo、Cascade、Centennial を対象に、アメリカ国内および国外産地の比較を行いました。Amarillo ではアメリカ・ワシントン州と他の産地の間に明確な違いがあり、Centennial でもベルギーとワシントン州の間に違いが確認されています。Cascade も産地間で差異は見られましたが、前二者ほど顕著ではなかったようです。残念ながらビール化実験は 1 回だけで、種々の制約があったのでしょう。もし差異が繰り返し観察されるのであれば、安定した共分散と解釈でき、普遍的な環境要因と遺伝子型の相互作用、言い換えるなら「ある品種の特定産地における品質」として理解できるはずです。
同じ品種でも産地によって全く異なることは、現役時代に実際に経験しています。海外産ホップの買い付けを手伝っていた時期、毎年秋にヨーロッパの産地を回って品質確認をしていました。チェコの品種 Saaz は、北部のザテツ地方で栽培されたものではおしとやかで繊細、まとまりの良い香気が特徴です。一方、南のモラビア地方で栽培すると性格は一変し、華やかな香りが顕在化します。これは毎年変わらず、例外がありませんでした。かなり強い共分散が発生していたのだと思います。安定したテロワールと呼べるでしょう。
ちなみに、先述の Budvar 社ではザテツ地方の Saaz しか使用していません。確かに Budvar のビールには華やかさよりも、落ち着いた品格が似合います。
環境による分散(VE)が安定している例を、Moreol ら(2020)が報告しています。23 のホップ系統および品種をアメリカ 2 か所で比較したところ、炭素数 6 の Hexyl 基を持つ配糖体がすべてのホップで高くなる産地があるというのです。品種という遺伝子型は関係なく、先の式における遺伝子型(VG)と遺伝子型と環境の共分散(COVGE)が消え、環境(VE)の分散しか残らないことになります。これも一種のテロワールといえます。
配糖体は水溶性のためルプリンではなく苞や葉に含まれています。ビール中では酵母によって糖部分が加水分解され、香気成分に変化します。Hexyl 配糖体の代表例は 1-hexanol で、青臭くビールには好ましくない香りとされています(Moreol ら 2020)。
なぜこのように一律に高くなるのでしょうか。論文では結論付けられていませんが、その役割を考えてみましょう。葉に含まれる Hexyl 配糖体は、昆虫の食害を受けると直ちに加水分解され、大気中に放出されます。このような香気成分は近年の研究によって「植物間コミュニケーション」を担うシグナルであることが示されています(有村ら 2019)。すなわち、食害を受けていることを周囲に知らせ、近傍の植物はそれを受けて防御機構を働かせるのです。具体的には、消化に悪いポリフェノールを生成したり、天敵を誘引して害虫を捕食してもらったりします。揮発性の香気成分を使い、捕食が起きている環境を操るのです(Boncan ら 2020)。
もし Hexyl 配糖体が防御機構の一つとして機能するのであれば、その産地には特定の昆虫やダニが発生しやすい環境があるのかもしれません。そのような特徴的な環境が、どのホップにも共通する普遍的な遺伝子(少なくとも品種改良で操作されていない遺伝子)の発現程度を制御していると考えられます。
テロワールの研究が先行しているワインについては、Anesi ら(2015)がイタリアのワイン用ブドウ品種 Corvina を対象に、7 か所で 3 年にわたる調査を行いました。その結果、特定産地において安定したテロワールが存在することを、遺伝子の発現、酵素などのたんぱく質合成、代謝産物レベルで証明しています。栽培環境と遺伝子型の共分散とは、環境が遺伝子やたんぱく質の発現に影響を与え、代謝系を変化させ、最終的に表現型を変化させている数値なのです。
以上が、テロワールの科学的側面に関する私の個人的な解釈です。
ノート4-1 遺伝学用語の解説
Repeatability(Falconer 1989)
「集団」の二カ年にわたる表現型値の変動(分散:VP)は、以下のように分けられます。
- 個体間の違い、すなわち遺伝的要因による分散(VG)
- 避けることのできない環境による分散(VEg)
- 二カ年の環境の違いによる分散(VEs)
Repeatability を r とおくと、次の式で表されます。
r =(VG + VEg)/ VP
VEs = 1 - r
表現型値のうち、どの程度が遺伝的に決まっているかを示すのは VG / VP です。したがって r ≧ VG / VP となります。Repeatability は表現型値の遺伝的部分の上限値、逆に言えば環境からの影響の下限値を示す指標なのです。
遺伝率
遺伝率は「子が親とどれだけ似ているか」を示す指標です。表現型値を測定する子供たちがどのような交配から生まれてきたかによって、計算方法が変わります。
村上(1999)は、子の母親への回帰係数(データは 1 年のみ)を、55 の子供ホップとそれぞれの計 11 母本との共分散から計算しました。
Henning と Townsend(2005)は、5 母本と 5 父本の総当たり交配を行い、1 家族内の子供たちとその母本・父本との共分散から遺伝率を求めました。計算の詳細や値の意味の違いは専門的になりすぎるため割愛しますが、Henning と Townsend(2005)の実験は大規模で、環境の影響を排除し誤差を小さくすることができました(5 母本 × 5 父本 = 25 家族、子供ホップ 5 個体/家族、4 反復/年、2 年間で 25 × 5 × 4 × 2 = 1000 試料)。
連鎖地図
染色体上で対を形成した遺伝子は、複製や減数分裂の際に組み換え(交叉とも呼びます)が起こり、遺伝子の対の片方からもう片方へ移動することがあります。交配を行うと、染色体上で距離の近い形質は一緒に分離するため、複数の形質が同時に観察されます。一方、組み換えが起こると複数の形質は独立して観察されます。
このように複数の形質が同時に観察されることを、遺伝学では「連鎖している」と呼びます。連鎖の程度は遺伝子間の距離として表され、組み換えが起こる頻度によって計算できます。多数の遺伝子をこの連鎖の程度に応じて地図化したものが「連鎖地図」です。
ノート4-2 成分比率の実験系の補足
仮に MURAKAMI SEVEN の雄が出現して自殖できたとしたら、話は大きく変わります。MURAKAMI SEVEN に MURAKAMI SEVEN を交配することになるのです。
自殖した子どもたちでは、MURAKAMI SEVEN が持つ遺伝子座のうちヘテロ型が分離し、それに伴って表現型も分離します。親の MURAKAMI SEVEN に類似性が高いものから低いものまで、子どもホップが連続的に現れるはずです。
雌ホップを雄に変えて交配するなど空想の話と思われるかもしれませんが、実は現実のものとなりそうなのです。2023 年、増殖中の畑に突然変異を起こし、雄に変わった MURAKAMI SEVEN が 1 個体、突如として現れました。驚きました。ホップの場合、ある年に性別が自然に変わる事例はこれまでも報告されています。私自身にとっては 2 度目の体験でしたが、原因は今のところ不明です。
花以外の外観では見分けがつかず、そっくりでした。ところが花が咲いたら雄だったのです。受精能力はないはずでしたが、自殖交配種子も相当量手に入りました。来春に種を蒔きますが、本当に発芽するかどうかは分かりません。うまくいけば詳細と結果を、いずれご報告できると思います。
引用文献
- Anesi, A., Stocchero, M., Santo, S. D., Commisso, M., Zenoni, S., Ceoldo, S., Tornielli, G. B., Siebert, T. E., Herderich, M., Pezzotti, M. and Guzzo, F., Towards a scientific interpretation of the terroir concept: plasticity of the grape berry metabolome, BMC Plant Biol., 15, 191, 2015
- Biendl, M., Engelhard, B., Foster, A., Gahr, A., Lutz, A., Mitter, W., Schmidt, R. and Schonberger, C., Hops Their cultivation, composition and usage, Fachverlag Hans Carl, Nuremberg, Germany, 2014
- Boncan, D. A. T., , Tsang, S. S. K., Li, C., Lee, I. H. T., Lam, H.-M,, Chan, T.-F. and Hui, J. H. L., Terpenes and Terpenoids in Plants: Interactions with Environment and Insects, Int. J. Mol. Sci., 21, 7382, 2020
- Burgess, A. H. Hops. Leonard Hill, I History of hop growing, London, 1964
- Falconer, D. S. Introduction to quantitative genetics. 3rd edition, Longman Scientific & Technical, New York, 1989
- Furukawa, J., Murakami, A. and Ichii, T., Preliminary results of a genetic approach for comparing aroma compounds, 2012年度 BCOJ年次大会要旨集, 21, 2012
- Henning, J. A. and Townsend, M. S., Field-based estimates of heritability and genetic correlations in hop, Crop Sci., 45, 1469-1475, 2005
- Holle, A. V., Muylle, H., Haesaert, G., Naudts, D., Keukeleire, D. D., Roldan-Ruiz, I. and Landschoot, A. V., Relevance of hop terroir for beer flavour, J. Inst. Brew., 127, 3, 238-247, 2021
- Hughes, P. S. and Simpson, W. J., Bitterness of Congeners and Stereoisomers of Hop-Derived Bitter Acids Found in Beer, J. Am. Soc. Brew., 54, 4, 234-237, 1996
- Hughes, P., The Significance of Iso-α-Acids for Beer Quality Cambridge Prize Paper, J. Inst. Brew., 106, 5, 271-276, 2000
- Koie, K., Inaba, A., Okada, Y., Kaneko, T. and Ito, K., Construction of the genetic linkage map and QTL analysis on hop (Humulus lupulus L.), Acta Horticulturae, 668, 668, 59-66, 2005
- McAdam, E. L., Freeman, J. S., Whittock, S. P., Buck, E. J., Jakse, J., Cerenak, A., Javornik, B., Kilian, A., Wang, C-H., Andersen, D., Vaillancourt, R. E., Carling, J., Beatson, R., Graham, L., Graham, D., Darby, P. and Koutoulis, A., Quantitative trait loci in hop (Humulus lupulus L.) reveal complex genetic architecture underlying variation in sex, yield and cone chemistry, BMC Genomics, 14, 360, 2013
- Morcol, T. B., Negrin, A., Matthews, P. D. and Kennell, E. J., Hop (Humulus lupulus L.) terroir has large effect on a glycosylated green leaf volatile but not on other aroma glycosides, Food Chem., 321, 15, 126644, 2020
- Neve, R. A. Hops. Chapman and Hall, London, 1991
- Okada, Y., Koie, K., Unaba, A., Kaneko, T. and Ito, K., Molecular markers for alpha acids; Study for practical application, International Hop Growers’ Convention, Proceeding of Sciental Commission, 56-59, 2007
- Patzak, J., Nesvadba, V., Krofta, K., Henychova, A., Marzoev, A. I. and Richard, K., Evaluation of genetic variability of wild hops(Humulus lupulus L.) in Canada and the Caucasus region by chemical and molecular methods, Genome, 53, 545-557, 2010
- Schönberger C. and Kostelecky, T., 125th anniversary review: the role of hops in brewing, J. Inst. Brew., 117, 3, 259-267. 2011
- Wang, G., Tian, L., Aziz, N., Broun, P., Dai, X., He, J., King, A., Zhao, O. X. and Dixon, R. A., Terpene Biosynthesis in Glandular Trichomes of Hop, Plant Physiol., 148, 3, 1254-1266, 2008
- 有村源一郎,上村卓矢,八代拓也, 植物アロマ成分を用いた有機栽培システムの開発とヒト健康増進効果への応用 植物アロマの潜在能力に注目!, 化学と生物, 57, 7, 428-432, 2019
- 飯島 陽子, 香辛料・ハーブとその香り~香気生成メカニズムとその蓄積, におい・かおり環境学会誌. 45, 2, 平成26年, 2014
- 富江弘幸, 教養としてのビール, SBクリエイティブ株式会社, 東京, 2019
目次
はじめに
第1章 ホップの苦味の科学
・はじめに抗菌性あり!
・α酸を貯蔵するルプリンの役割
・ホップの抗菌性
・抗菌性のメカニズム
・ホップの苦味成分たち
・Cohumulone論争
・苦味成分の脇役たち
・フルーツビールの教え
・苦味価とは
・ノート1-1 毬花
・ノート1-2 pH
・ノート1-3 成分の性質
・ノ-ト1-4 分析法について
・引用文献
第2章 ホップの香りの科学
・香り成分分析技術の進歩のおかげ
・ホップとビールでの成分組成の違い
・ホップ香気成分の世界
・香気成分の間に起こる相互作用
・香りはどうように感知されるのか
・香気成分データの解析
・ホップ品種の香気のアイデンティティーはどの様に決まる?
・ノート2-1 テルペン類
・ノート2-2 濃度単位のお話し
・ノート2-3 含硫成分の分析
・ノート2-4 多変量解析
・引用文献
第3章 ホップ使用技術
・ビールの作り方
・苦味の付け方
・ホップ香気の付け方
・ホップの加工・調製品
・生ホップの特徴とは
・ルプリンパウダーの教え
・引用文献
第4章 ホップの育種
・交配育種の始まり
・雄ホップと交配育種
・遺伝の基礎
・ホップの遺伝学
・Cohumulone(Coh)論争の考察
・新ホップ品種MURAKAMI SEVENのいきさつ
・ホップ品種のアイデンティティーとは
・ホップのテロワール
・ノート4-1 遺伝学用語の解説
・ノート4-2 成分比率の実験系の補足
・引用文献
第5章 ホップの進化遺伝学
・分子進化遺伝学
・PCRの登場
・分子進化時計
・野生ホップ収集開始
・世界の野生ホップのDNA解析
・Humulus属の構成
・DNA塩基の違い
・ホップの起源の地
・分岐年代の推定
・ホップの伝播のシナリオ
・マイクロサテライトDNA多型
・コーカサス地方のホップ
・日本の野生ホップ
・日本野生ホップ、カラハナソウの分布
・カラハナソウと氷河期(最終氷期)との関係
・ノート5-1 分子進化時計のキャリブレーション
・ノート5-2 マイクロサテライトDNA多型
・ノート5-3 コーカサス集団とヨーロッパ集団の分岐年代推定
・引用文献
第6章 ホップを取り巻く環境の変化
・ビール事情の変化
・世界のホップ育種事情の変化
・世界のホップ生産事情の変化
・日本のホップ事情
・遠野、Now!
・ホップの新たな可能性-ホップの健康機能性-
・引用文献
付録:ホップの育て方
・成長のフェーズ
・ホップの育て方-作業-
・ホップの育て方-施肥-
・ホップの育て方-病虫害、自然発生被害-
・ホップの収穫
・ホップ栽培、あるある!
・将来のホップ栽培のために
・引用文献
おわりに
※校正しながら連載を進めますので、目次の一部が変更となる可能性があります
お知らせ
連載の更新情報は、GOOD HOPS公式SNS(X・Facebook)で随時お知らせします。
ぜひフォローしてチェックしてください。
X
facebook
また、村上が手がけたGOOD HOPSのビールも現在販売中です。下記のリンクよりご確認ください。
一般の方向けオンラインショップ
業務店さま向けオンラインショップ


お問い合わせ
施設やTAPROOMに関するご質問、自社ビールのお取り扱いに関するお問い合わせ(小売店様・飲食店様向け)、取材のご依頼など、どうぞお気軽にご連絡ください。
スタッフ一同、美味しいビールとともにお待ちしております!
